SISTEM REPRODUKSI IKAN BETINA : Anatomi dan Fisiologisnya
Reproduksi – pada ikan adalah biseksual, hermafrodit atau partenogenik. Proses yang paling mendominasi adalah reproduksi biseksual. Pada ikan seperti itu, jenis kelaminnya terpisah, mis., Individu dioecious. Dalam beberapa ikan, kedua jenis kelamin hadir dalam individu yang sama, yaitu, ikan tersebut adalah hermafrodit.
Contohnya adalah Perca, Stizostedion, Micropterus. Pada beberapa ikan, hermafroditisme remaja telah diketahui. Pada Poe-cilia Formosa terjadi partenogenesis, sebenarnya prosesnya adalah gininesis, yaitu, perkembangan anak muda tanpa pembuahan.
Gonad dari elasmobranch dikembangkan oleh migrasi sel benih primordial dari endoderm ekstra-embrionik yang berasal dari epitel peritoneum dan analog antar ginjal. Dalam teleosts juga sel-sel benih primordial berasal dari punggung genital (seperti pembengkakan). Sel benih primordial cukup besar dan bentuknya agak oval dengan nukleus bulat besar yang mengandung satu nukleolus besar.
Punggung genital berkembang menjadi korteks dan medula pada vertebrata. Tetapi gonad pada teleost dan cyclostoma hanya terdiri dari korteks dan tidak memiliki jaringan medula.
Pada elasmobranch berkembang menjadi medula, tetapi pada wanita mereka berkembang menjadi korteks. Kemudian sel-sel benih primordial mengalami meiosis dan berkembang biak dan akhirnya berdiferensiasi menjadi gonad jantan dan betina.
Ikan memiliki sepasang gonad bilateral. Mereka umumnya simetris. Mereka ditangguhkan dari bagian dorsal rongga tubuh oleh mesenteries melintasi atap rongga tubuh dalam hubungan erat dengan ginjal.
Mesenterium pada wanita disebut mesovarium. Mereka juga ada di Neoceratodus. Sedangkan pada pria dikenal sebagai mesorchium. Ia kaya didukung oleh pembuluh darah dan serabut saraf.
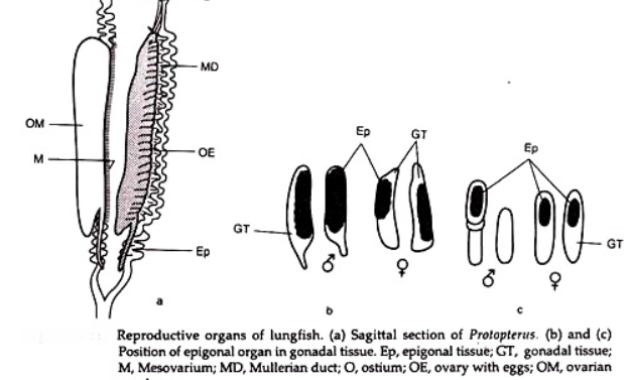
Organ Epigonal Hematopoietik
Dalam semua elasmobranch gonad dikaitkan dengan organ epigonal hemopoietik. Testis seperti ovarium juga berhubungan dengan organ epigonal. Mereka meluas ke kloaka, mungkin relatif kecil di Heptanchus atau tidak ada di Squalus. Dalam Scyllium, testis menyatu dengan organ epigonal.
Dipasangkan dalam Cetorhinus, salah satu sisi kanan berbaring posterior dan sedikit dorsal ke ovarium dan ditangguhkan oleh ekstensi posterior mesovarium. Panjangnya sekitar 60 cm dan diameternya 20 cm. Daerah anterior secara intim menyatu dengan sepertiga posterior ovarium.
Organ epigonal kiri pada wanita memiliki ukuran dan bentuk yang mirip meskipun tidak terkait dengan gonad. Pada hiu betina dari banyak genus, hanya satu ovarium yang berkembang (Scyliorhinus, Pristiurus, Galeus, Mustelus) dan, oleh karena itu, sesuai dengan organ eipgonal betina sering tidak berpasangan, sehingga lebih baik berkembang di satu sisi daripada yang lain.
Organ Reproduksi Betina
Organ reproduksi betina terdiri dari ovarium, saluran telur dan pada beberapa ikan papila pseudo-sanggama. Ovarium dalam kaitannya dengan saluran telur dan transportasi sel telur dibedakan menjadi tipe sitovarian, tipe semicytovarian, dan tipe gymnovarian. Pada tipe cytovarian, lumen ovarium dihubungkan dengan saluran telur dan dilepaskan dalam air melalui pori genital.
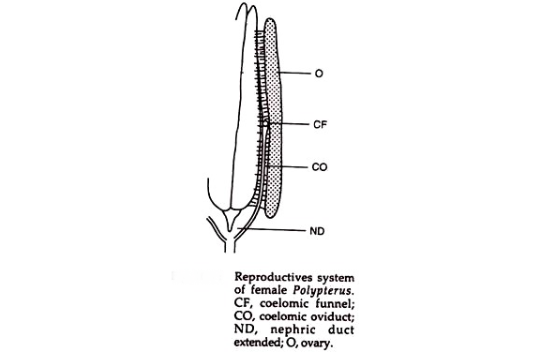
Dalam jenis semicytovarian, oosit yang menggantikan saluran telur melewati alur transparan berbentuk corong, yang membuka ke pori genital. Kondisi seperti ini terdapat di Notopteridae, Osteoglassidae, dll., Di mana saluran telur mengalami degenerasi sebagian atau seluruhnya dan, oleh karena itu, sel telur dimasukkan ke dalam rongga coelomic dan kemudian dibawa melalui pori-pori atau corong.
Jenis indung telur ovarium tidak kontinu dengan saluran telur. Ovarium menggantung seperti tirai dan oosit diovulasi ke dalam rongga selom atau peritoneum dan dibawa ke saluran telur oleh silia.
Biasanya ovarium berbentuk seperti kantung dan berpasangan dan struktur simetris baik di teleosts dan elasmobranchs dan fungsional. Namun, pada beberapa elasmobranch, ovarium kanan menjadi fungsional sementara pada beberapa kasus ovarium kiri hanya berkembang.
Histologi
Ovarium mengandung jaringan pendukung yang dikenal sebagai stroma, yang mengandung oogonia dan oosit. Oogonium (sel germinal) setelah multiplikasi berkembang menjadi oosit primer. Sel oogonium pada tahap awal pengembangan adalah sel besar dengan nukleus besar yang memiliki nukleolus yang menonjol dengan sitoplasma kromo-fob. Ukuran sel menjadi lebih kecil setelah multiplikasi, tetapi tidak penting.
Perbedaan penting adalah pada kromosom. Kromosom menjadi benang seperti pada nukleus (tahap leptoten). Kromosom menjalani tahap zygotene diikuti oleh tahap pachytene dan akhirnya diplotene. Pada tahap perkembangan ini hingga oosit dikelilingi oleh sel-sel folikel. Sel-sel folikel multilayer dalam elasmobranch dan amniota tetapi lapisan tunggal pada teleost.
Pada mamalia, sel-sel folikel menimbulkan epitel bertingkat, stratum granulosa yang mengelilingi oosit dan akhirnya berkembang biak untuk membentuk sel-sel granular. Sel-sel stroma segera mengelilingi folikel, membentuk selubung sel, folikel teka yang dibedakan menjadi teka eksterna dan teka interna.
Dalam elasmobranch, theca interna terletak tepat di bawah eksterna dan terdiri dari 4 hingga 6 baris sel rata dan memanjang, yang dindingnya diperluas ke luar menuju proses (Dodd, 1983). Sel theca externa berfungsi sebagai sekretaris dan mengandung organel seperti ER dan mitokondria.
Ini mengandung fibroblas, serat kolagen dan kapiler darah serta sel penghasil steroid. Oosit yang matang memiliki dua lapisan, lapisan teka luar dan lapisan granulosa dalam, keduanya dipisahkan satu sama lain oleh membran dasar yang berbeda.
Ovarium mengandung banyak oosit dalam berbagai tahap perkembangan dan degenerasi. Atresia folikuler adalah proses degeneratif di mana oosit dalam berbagai tahap pertumbuhan dan diferensiasi mereka hilang dari ovarium ikan dan ovulasi. Folikel kehilangan pertumbuhan dan diferensiasi mereka. Mereka tidak mensintesis hormon steroid dan menjalankan fungsi endokrin apa pun.
Folikel dengerasi terdiri dari dua jenis, korpora lutea dan corpora atretica. Pada korpora lutea, oosit tertutup dikeluarkan dari folikel oleh sel granulosa yang menyerang dan menanamkan sel teka. Corpora lutea dan corpora atretica hadir di ovarium holocephalon.
Dalam spesies ovipar dan vivipar mereka biasanya hadir pada akhir musim panas. Mereka mungkin mewakili oosit yolky yang karena beberapa alasan telah gagal diovulasi. Dalam Squalus acanthias corpora atretica tidak ada di ovarium.
Vitellogenesis
Telur pada awalnya tanpa kuning telur tetapi vitellogenesis terjadi kemudian (atresia). Pada vertebrata ovipar dan ovovivipar telah lama diketahui bahwa darah wanita dewasa mengandung lipofosfoprotein pengikat kalsium, vitellogenin, yang disintesis di hati dengan stimulasi steroid jenis kelamin wanita.
Ini berpindah dari plasma ke oosit di mana ia memunculkan protein kuning telur, lipovitelin dan phosvitin (Wallace, 1978). Vitellogenesis pada dogfish mirip dengan yang ditemukan pada vertebrata lain. Craik (1978) mengukur laju di mana vitellogenin disintesis dan diubah menjadi butiran kuning telur.
The vitellogenesis berlangsung, di mana akumulasi zat kuning telur terjadi. Perubahan mencolok yang terlihat adalah sitoplasma yang sebelumnya bersifat basofilik, sekarang menjadi asidofilik. Ada tiga substansi kuning penting — vesikel kuning telur, butiran kuning telur, dan tetesan minyak.
Vesikula kuning telur disediakan dengan glikoprotein yang diwarnai sedikit merah dengan eosin tetapi PAS positif. Mereka mengambil noda merah tua. Vesikula kuning telur kemudian menjadi alveoli kortikal dan mengambil bagian dalam pembentukan ruang perivitelline.
Gumpalan kuning telur terutama terdiri dari lipoprotein dengan beberapa karbohidrat dan zat lain. Mereka lemah positif terhadap PAS. Tetesan lama umumnya mengandung gliserida dan sejumlah kecil kolesterol. Dengan selesainya gerakan vitellogenesis vesikel germinal, fusi butiran kuning telur dan pengelompokan tetesan minyak terjadi.
Setelah pergerakan vesikula germinal ke kutub hewan, pembelahan meiosis pertama terjadi dan tubuh kutub pertama dilepaskan.
Selanjutnya divisi meiotik kedua dimulai dan telur di mana divisi ditangkap pada metafase diovulasi. Chieffi; percaya bahwa corpora lutea sejati (yaitu, dalam arti fungsional mamalia) ditemukan dalam elasmobranch yang ovipar dan vivipar.
Dalam yang pertama (S. stellaris) mereka diyakini berkembang dari folikel yang mengalami ovulasi (corpora lutea) dan yang terakhir (T. marmorata) oleh atresia folikular (corpora atretica).
Steroid hadir di ovarium elasmobranch. 17 beta estradiol dan jejak progesteron dan estrogen ada di ovarium Squalus. Hal ini ditunjukkan dalam Squalus acanthias bahwa progesteron dapat disintesis dari (HC) pregnenolon.
Enzim yang diperlukan untuk steroidogenesis dalam ovarium Squalus acanthias seperti glukosa-6 fosfat dehidrogenase (G-6PDH), 3 beta-hydroxysteroid dehydrogenase (3 beta HSD), 3 alpha-hydroxysteroid dehydrogenase (17 beta HSD) dan 20 beta hydroxysteroid dehydrogenase (17 beta HSD) (20 beta HSD) hadir di ovarium.
3 beta HSD ditemukan di corpora lutea tetapi tidak di corpora atretica dan 3a-HSD lemah hadir di granulosa dari corpora atretica.